| - Home page |
| - No.234 目次 |
| + 研究紹介 |
| - ISAS事情 |
| - M-V事情 |
| - 宇宙を探る |
| - 東奔西走 |
| - 惑星探査のテクノロジー |
| - いも焼酎 |
| - 編集後記 |
| - BackNumber |
| 用語解説 |
| 極限環境微生物 |
進化と酵素と好熱菌と
東京薬科大学生命科学部 山 岸 明 彦
私は好熱菌や超好熱菌などの極限環境微生物を実験材料として,大きく分けて二つの側面から研究しています。その第一は生命の初期進化に関する研究で,もう一つは蛋白質工学に関する研究です。細かいテーマも含めるとかなり多くなりますが,その中から何か一つでも興味を持っていただければと思い,それらを含めて紹介させていただきたいとおもいます。お忙しい方は,小見出しだけご覧下さい。
I.生命の初期進化の研究(参考文献 1ー5)
1.全生物の共通の祖先はどんな生き物だったか
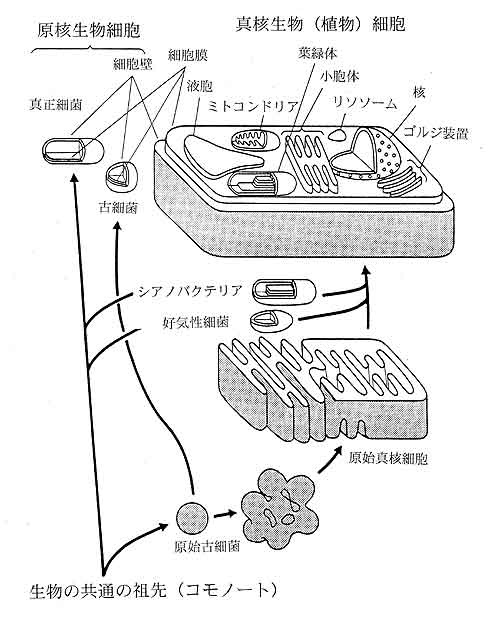
生命は今から約40億年前に誕生したと考えられています。地球上の温泉や海底の熱水噴出口に棲む現存の超好熱菌や好熱菌を調べることから,40億年前の生物がどのような生物であったのかが分かるのではないかと思っています。図1は,現在最も信頼されている全生物の系統樹を示しています。系統樹の根元は生命の起源を表し,系統樹の枝の先端が現在の様々な生物種に対応しています。現存する生物は大きく3つに分類されています。それらは,真正細菌(いわゆる良く知られたバクテリア)と古細菌(極限環境にすむ特殊な原核生物),それに真核生物(動物,植物,カビ,原生動物など)です。生命の起源の後,図1でコモノートと書いてある点で約38億年前に生命は真正細菌と古細菌に分かれました。古細菌の様々な性質を調べて,真正細菌と比較することから両者の(従って全生物の)共通の祖先がどのような生物だったのかが分かるのではないかと考えています。
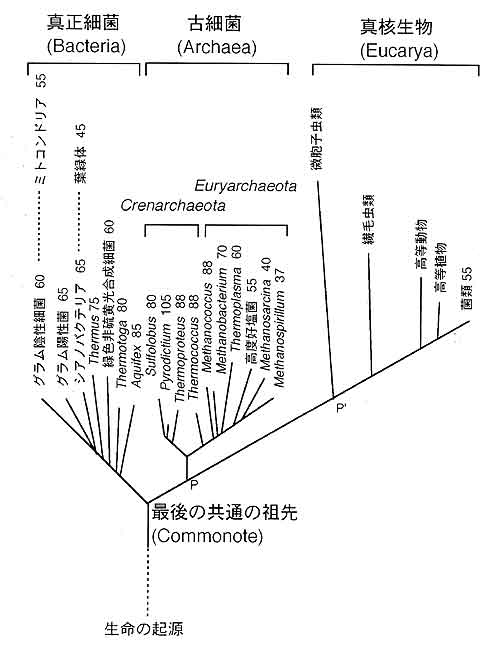
図1 全生物の進化系統樹。生物界は三つに分けられている。
真核生物のミトコンドリアと葉緑体はそれぞれ専用のrRNA遺伝子
を持ち,その配列を解析すると,真正細菌の中に分類される。