No.224
1999.11 |
<研究紹介>
ISASニュース 1999.11 No.224
|
鞭毛・繊毛の世界  運動制御から見た生殖と走性
運動制御から見た生殖と走性
東京大学大学院総合文化研究科 奥野 誠
生殖と鞭毛・繊毛運動が私の研究室のキーワードです。生物の生殖は多種多様です。多くの動物では,形態も機能も大変異なった二種類の配偶子,すなわち卵と精子を形成し,その融合(受精)によって新しい個体が誕生します。大きくてタンパク質などに富んだ卵と,小さくて活発に動きまわる精子という組み合わせは,進化の過程が生んだ合理的な戦略であるとされています。この精子を動かしているのが微小管を骨格とする構造すなわち鞭毛です。微小管系は,筋肉などを構築する微小繊維系と並んで,細胞運動の二本柱の一つです。そして微小管系の代表的なものが鞭毛・繊毛で,一般的に直径が24nmの微小管が二本結合したようなダブレット微小管9本を周上に配し,中央に2本のシングレット微小管をもつ“軸糸”を骨格とします(図1)。多くの動物の精子や,原生生物では,この直径約0.2μmの軸糸が鞭毛や繊毛そのものですが,哺乳類の精子ではこの軸糸の外側に外側粗大繊維などのアクセサリーファイバーをもっており,鞭毛は太くなっています。また軸糸は運動器官のみならず,平衡感覚や聴覚において感覚繊毛として働いています。ところでこれらの鞭毛・繊毛は真核生物に特有の器官であり,バクテリアなど原核生物にある鞭毛とは構造も運動の仕組みも全く異なるものです。
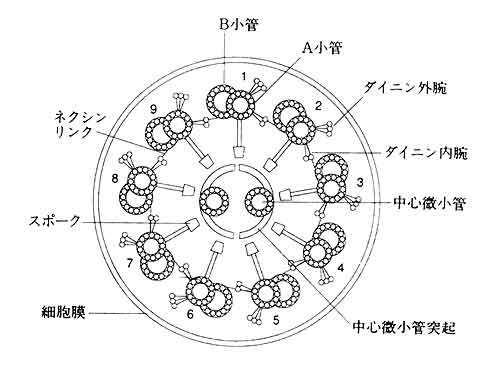
図1.鞭毛(クラミドモナス)の断面の模式図(毛利1976を改変)。9+2構造を示す。
現在進行中の研究テーマは,鞭毛や繊毛がどのような仕組みで屈曲波を作りだしているのか。その運動の制御はどのような仕組みによっているのか。そしてそのような制御機構はどのようにして構築されていくのかということです。材料としては精子及び繊毛をもつニハイチュウなどを用いています。次に幾つかの研究結果を紹介したいと思います。
(1) 精子の運動制御機構
鞭毛・繊毛において運動の力を発生するのは軸糸の部分であり,その他の構造,すなわち哺乳類精子がもつような外側粗大繊維,繊維鞘といったものの生理学的意義ははっきりしていません。軸糸の9本のダブレット微小管は“腕”が発生する力によって能動的滑りを起こします。ダブレット微小管の頭部側の末端は中心体で固定されているため,微小管のずれは屈曲に変換されます。9本のダブレット微小管のほぼ全長にわたって規則的に存在する“腕”が,時間的,空間的規則性をもって滑り力を発生するために,鞭毛の規則的な屈曲振動が生じると考えられています。
(1.1)魚類精子の運動開始機構
ところで精子は精巣で作られた後,すぐに動き始めるわけではなく,生殖環境が整った時点で運動を開始します。体外受精の魚類などでは非常にはっきりしており,多くの場合放精と同時に運動を開始します。たとえば淡水魚精子では,一般に体内より低浸透圧に曝されると,すなわち淡水中に放精されると,それを刺激として運動を開始します。このような細胞外からのシグナルが細胞膜を介して細胞内情報に変換され,それが鞭毛軸糸の屈曲を引き起こすと考えられます。ところでサケ科魚類の場合は,放精によって細胞外カリウムイオン濃度が低下することが引き金となります。私たちはシロサケやニジマス精子を用いて,この細胞外の引き金が細胞内の情報伝達物質であるcAMP合成を誘導し,それが鞭毛内の15kDaタンパク質のリン酸化を引き起こすことが鞭毛軸糸の活性化,ひいては鞭毛の屈曲運動を開始させることをつきとめました。
(1.2)哺乳類精子の運動調節
哺乳類精子の運動はもっと複雑です。精巣で完成した精子は,運動能はほとんどもっていません。精巣上体を通過して始めて運動能が付与され,精嚢,前立腺などからの分泌液に希釈されると活発な運動を開始します。さらに雌性生殖器内で受精能獲得と呼ばれるプロセスを経て初めて受精が可能になります。受精能獲得の過程で,鞭毛は超活性化と呼ばれるタイプの運動を呈することになります。これらの一連の現象を解明することは,この過程の生殖における意義を明らかにするためのみならず,鞭毛運動の制御機構を知る上でも大変有意義なことです。
我々はマウスやハムスターを用いて,哺乳類精子の運動能獲得と運動調節の機構を調べてきました。成熟したマウス精子は重炭酸イオンによって運動が活性化します。この時,細胞内では65kDaのタンパク質がリン酸化されます。そして同時に微小管間の相互作用が変化し,微小管の滑り速度が上昇します。図2は,細胞膜を除去した精子において,ダブレット微小管が滑り出してループを形成しているところです。矢尻で示された繊維鞘の部分に注目すると,活性化する前の精子と後の精子では異なった反応を示していることがわかります。微小管の滑り速度もおよそ2倍に増加していました。同様のことがカルシウムで活性化したハムスター精子でも観察されました。こちらでは36kDaのタンパク質がリン酸化されます。またこのタンパク質に対する抗体を作用させると,活性化の作用が押さえられることもわかりました。このように,精子の運動活性化には精子鞭毛のタンパク質のリン酸化が重要な役割を演じていることがわかりました。
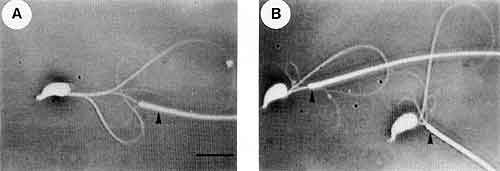
図2.細胞膜を除去したマウス精子で微小管が滑り出してループを作っている様子。
A:活性化していない精子。B:活性化した精子。矢尻は繊維鞘を示す。黒棒は10μm。
受精能獲得過程を経ると鞭毛の運動も大きく変わり,超活性化運動をするようになります。図3はそれを示しています。そしてこの変化においても今度は80kDaタンパク質のリン酸化が起こることがわかりました。
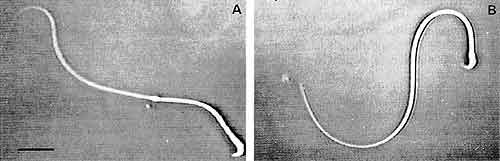
図3.活性化精子(A)と受精能獲得し超活性化した精子(B)。黒棒は25μm。
このように,運動の変化に伴い,ここでは紹介しなかったものも含めて,様々なタンパク質のリン酸化が起こります。我々は精子の運動の一連の変化の仕組みを,タンパク質のリン酸化を軸にして解明したいと考えています。
(1.3)生殖と重力環境
生殖は非常にデリケートなものです。環境ホルモンはそのよい例で,わずかなホルモン用物質が性転換を引き起こしたり,生殖能を失わせることがわかってきました。我々は環境と雄の生殖という視点から,地球環境,特に重力環境が生殖にどのような影響を与えるかを調べてきました。
実は精子の運動そのものも重力による影響を受ける可能性があります。精子を遠心分離機にかけると,運動性の高いものが早く沈降するという経験的事実があります。1Gでも,哺乳類精子で調べたところ泳ぐものでは運動をしておらず自然沈降するものに比べると早く沈降することがわかりました。ただし交尾をする哺乳類ではあまり重要な違いでないと思われます。
さてマウスの雄では,誕生後精巣は発達し続け,およそ5週齢で完成された精子が精巣上体に出現します。低重力環境でこのように長期間飼育することは地上では無理です。そこで,遠心飼育機による加重実験によって過重力の影響を調べ,微小重力の影響を推測することにしました。その結果,驚いたことに,加重によって精子形成は抑制されるどころかむしろ促進されるのではないかという結果を得ました(図4)。3Gで遠心飼育を行ったところ,精巣重量はあまり増えないのですが体重は更に抑制されます。その結果,精巣重量/体重の値をとると,1Gのものとは歴然とした差がみられました。しかし精子の性状ではほとんど差がないことがわかります。飼育条件を4Gにすると発育が非常に悪くなること,多世代にわたる飼育では出産数はあまり変わらぬものの大人になる個体が著しく減ることなど,尚多くの課題が残されてはいますが,大変興味深い現象です。微小重力ではどうか。これは現在是非調べてみたい問題です。
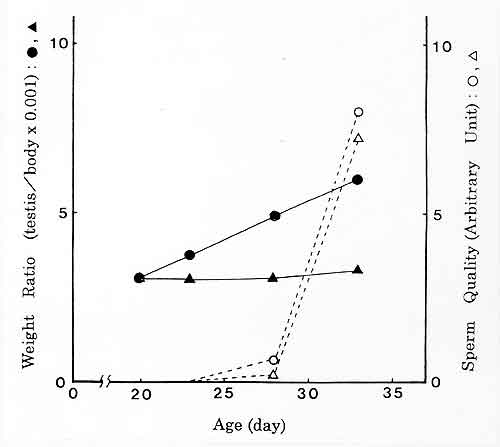
図4.過重力下での精巣の発達と精子性状。
実線は精巣重量と体重の比率,破線は精巣上体尾部から得た精子の性状の相対評価。
●,○は3G実験群,▲,△は1Gコントロール群。
(2) 重力走性と繊毛運動の制御
繊毛で運動する動物には単細胞の原生動物からより複雑な体制をもつものまであります。これらは走化性や走温性など様々な走性を示しますが,ゾウリムシなどでは重力走性を示すことが古くから知られています。精子とは違った意味で,繊毛運動の制御という点から大変興味深い問題です。
我々はニハイチュウという数十個の細胞のみで出来ている生物の滴虫型幼生が,強い正の重力走性を示すことを見いだし,その性質について調べています。この幼生は,先端にある頂端細胞が高密度な屈光体をもつため,頭部を下に向ける正の重力走性を示します。しかし,図5に示すように,下方へ泳ぐ遊泳速度は,水平方向の遊泳速度に自然沈降を加えた値より遙かに速いものでした。また遊泳方向を重力方向へ変えていく回転速度も,予測される値よりも速いものでした。これらの結果は,物理的原因に加えて,それを増幅する生物学的な仕組みが働いている可能性を示唆するものです。この生物学的な仕組みを明らかにしていきたいと考えています。
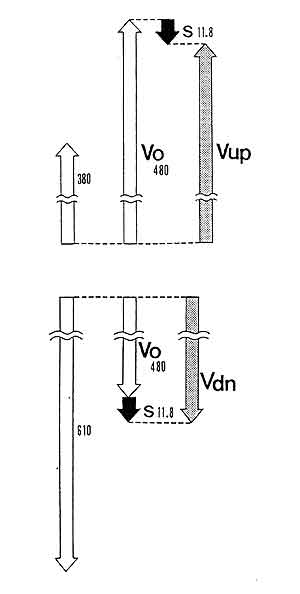
図5.ニハイチュウ滴虫型幼生の遊泳速度。
左端は上方及び下方への遊泳速度。中央は水平方向の遊泳速度(Vo)及び自然落下の速度(S)。
右端は水平方向の遊泳速度と自由落下の速度から推定される上方及び下方への遊泳速度。
数字はμm/s
結語
生殖も運動もきわめて生物学的な現象です。その両者を結びつけているものが鞭毛・繊毛であると言ってもよいかと思います。鞭毛・繊毛運動の制御機構を明らかにすることを通じて,精子の機能や細胞運動を明らかにしていきたいと考えています。
(おくの・まこと)
ISASニュース No.224(無断転載不可)
